如何判斷昆蟲的翅脈那條是那條
- 一、前言:
- 翅脈是昆蟲翅膀上的紋路,作為支撐翅膀的功能,依據延伸方向分為縱脈與橫脈,而昆蟲主翅脈通常為縱脈 ,在主翅脈內有氣管、血管及神經纖維。翅脈並非為平整的結構,而是連續的相接結構組合而成, 可以想成許多凸與凹兩種形式相接而成的結構,而每種昆蟲的翅脈分布不盡相同, 但彼此間有相似性 。
- 二、正文:
- 翅脈形式可作為分類昆蟲的重要依據,依據顏色或形式的不同而可能分出不同類群的昆蟲,再配合翅膀結構的差異可分為鞘翅目、鱗翅目、革翅目及雙翅目等等 。 如上述所述,每類昆蟲的翅脈結構可能不同,但是現今昆蟲翅膀的演化應當為單系群,也就是說所有有翅昆蟲具有相同的共同祖先,而有學者想要追溯原始昆蟲翅脈的結構 。
- Archedictyon,就是假想原始有翅昆蟲的祖先,Archedictyon的翅脈成為往後有翅昆蟲演化的模板,有翅昆蟲翅脈結構皆是由此逐漸演化成現今的模式 。 依據化石證據與學者推測,Archedictyon應當有6~8條縱脈,在1980年代昆蟲學者John Comstock和George Needham創造出假想的昆蟲翅脈模式圖,並命名各翅脈,此系統稱為Comstock-Needham System,現今大部分昆蟲學者仍沿用此假說(Meyer, John R. (5 January 2007)) [1] 。
- 以下將介紹各主要翅脈 :
- (一)縱脈 (Longitudinal veins) :
- 1.前緣脈(Costa,C):第一縱脈,為翅膀前緣
- 2.亞前緣脈(Subcosta,SC):第二縱脈(前緣脈之後),SC1~2
- 3.徑脈(Radius,R):第三縱脈,1~5條分支到翅緣(wing margin),R1(anterior radius)~5(The first posterior branch,fork radius sector,RS)
- 4.中脈(Media,M):第四縱脈,1~4條分支到翅緣(wing margin),M1~4
- 5.肘脈(Cubitus,CU):第五縱脈,1~3條分支到翅脈(wing margin),CU1(有時分支成CU1a與CU1b)~2
- 6.臀脈(Anal,A):在肘脈之後的未分支脈(unbranched veins),A1~4(unbranched anal veins)。
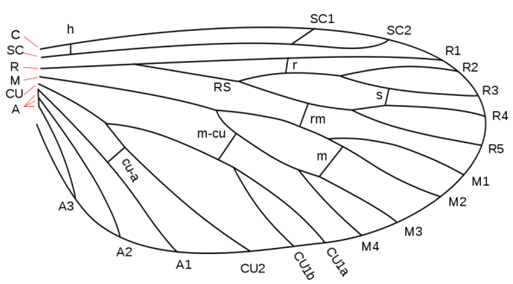
- 圖一、上圖為Comstock-Needham System的翅脈模式圖(Figure from: http://insects.about.com/[2])
- 大部分昆蟲的第一條縱脈為前緣脈 (Costa),但某些物種在前緣脈之前有Precosta (PC)結構,而目前在多數昆蟲觀察不到Precosta的原因為:Precosta與前緣脈融合為一體,因此只觀察到前緣脈的結構 (Gullan, P. J. and Cranston, P. S. (2004))[3] 。
- (二)橫脈 (Cross veins):橫脈通常與縱脈交錯,形成網狀結構
- 1.C-SC: 為前緣脈與亞前緣脈交錯
- 2.R: 鄰近分支徑脈彼此交錯
- 3.R-S: 亞前緣脈與徑脈交錯
- 4.R-M: 徑脈與中脈交錯
- 5.M: 鄰近分支中脈彼此交錯
- 6.M-CU: 中脈與肘脈交錯
- 7.CU-A: 肘脈與臀脈交錯。 (Data from:Debbie Hadley, March 17, 2015)[2]
- 雖然某些昆蟲,例如蜻蜓目(Odonata)有橫脈結構,但是許多昆蟲卻缺乏或是在演化過程中失去橫脈結構,橫脈的存在與縱脈交錯,提供翅膀較多的支撐力,如此可以使飛行能力更穩定或更快速。 依據化石證據與推測下,學者認為Archedictyon的飛行模式應該是前後翅各自拍動,然而以空氣力學的角度來看,這樣的運作模式是低效率的;高效率的運作模式應為前後翅同時拍動,目前有些物種的翅膀已經演化出此模式,而前翅的jugum(jugal area)參與其中的運作關鍵,jugum為某些昆蟲前翅與後翅重疊的部位,其功能為在飛行時將前後翅固定,並促使兩者能同時拍動,以達到高效率的飛行模式。
- 三、結論:
- 不同類群昆蟲的翅脈結構不盡相同,但是具有同源性,可追溯到原始有翅昆蟲Archedictyon,其翅脈作為往後有翅昆蟲的演化模板,而此假說由John Comstock和George Needham正式提出 。 雖然至今已歷經兩億年的演化過程,但是各昆蟲的翅脈結構依然具有相似性。翅脈結構可大致分為縱脈與橫脈,主要作為支持翅膀運作的功能,內部具有血管、氣管及神經等組織,提供昆蟲在飛行時所消耗的大量養分與氧氣,雖然某些昆蟲缺乏橫脈結構,但是依然保有飛行的功能 。從空氣力學看來,現今昆蟲以前後翅同時拍動以達到飛行目的的效率較原始有翅昆蟲高 。
- 四、參考資料:
- [1] Meyer, John R. (5 January 2007). "External Anatomy: WINGS". Department of Entomology, North Carolina State University. Retrieved 2011-03-21.
- [2]Debbie Hadley, March 17, 2015. Insect Wing Venation Diagram.about education. Retrieved March,22,2016,from http://insects.about.com/)
- [3]Gullan, P. J. and Cranston, P. S. (2004). The insects: an outline of entomology. UK: Blackwell Publishing. ISBN 1-4051-1113-5.
andrewneteye4343 發表在 痞客邦 留言(0) 人氣()
昆蟲翅的演化起源
- 一、前言:
- 昆蟲翅膀是由外骨骼突出生長的飛翔結構,通常昆蟲成體才具有飛行的能力,翅膀位於昆蟲的第二胸節(mesothorax)與第三胸節(metathorax),在具兩對翅的昆蟲中,兩對翅分別稱為前翅(forewings)與後翅(hindwings),而單翅昆蟲的後翅通常演化為平衡棍。即便知道翅膀目前的形態,但是目前仍沒有統一的理論解釋昆蟲翅膀的演化過程與起源。
- 二、正文:
- 目前對昆蟲翅膀起源的著名假說有三個,分別為 Paranotal hypothesis(胸部背板衍生假說)、Epicoxal hypothesis(腹部氣管式鰓演化假說)及 Endite-Exite Hypothesis(腿節演化假說)。
- (一)Paranotal hypothesis:
- 首先,在1976年Wootton認為具原始翅膀的昆蟲應該具有突起物(lobe),且該結構缺乏關節,當時應該是作為昆蟲從高處掉落時的緩衝結構,且隨時間演化在lobe中出現神經、血液及肌肉,逐漸演化為更具功能性的結構,從緩稱功能、滑翔功能到主動飛行功能(Wootton, RJ. 1976.)[1]。而在2010年 Gullan, PJ & Cranston, PS 認為原始翅膀應為胸部背板上的衍生結構(Gullan, PJ & Cranston, PS. 2010.)[2]。
- 支持這項假說的現象為大部分昆蟲具有往枝條移動的功能,在過程中難免會因為風吹或是其它自然因素而被迫掉落,當時具有lobe的昆蟲便有較高的機會能夠存活或更有利生存,這樣的性狀就在演化過程中被保留下來。但是仍然有反對者提出質疑,1985年Quartau認為從滑翔式翅膀演化為主動飛行翅膀是不太可能的,Quartau主張原始翅膀的功能應當就是主動飛行用。而現今因為缺乏翅膀上體節與肌肉的化石證據,所以Wootton的胸部背板演化假說遭到質疑(Grimaldi, David; Engel, Michael S. 2005.)[3]。
- (二) Epicoxal hypothesis:
- 在1906年Woodworth認為昆蟲翅膀由可移動的腹部鰓蓋(gill-cover) 或是氣管性質的鰓(tracheated gill)演化而成(Woodworth, CW. 1906.)[4]。在氣管鰓中有肌肉組織,且有小型的翼狀構造(winglets) ,隨著擺動可以不斷進行呼吸運動(Grimaldi, David; Engel, Michael S. (2005)[3]。此假說主張氣管鰓隨著演化,從呼吸結構逐漸轉為運動結構,最後形成現今的昆蟲翅膀。
- 鰓被認為是翅膀起源附肢結構的原因為鰓與翅膀同樣具有關節(joints) 與肌肉,且在某些物種幼蟲時期,而鰓外型像微型的翅膀。Michalis Averof認為若鰓與翅膀確實有相關性,則兩結構間的基因應當相似,為了驗證此推測,學者比較甲殼動物與蒼蠅(甲殼類為昆蟲近親,且使用鰓呼吸) 。藉由PCR研究兩者的DNA,學者在豐年蝦(brine shrimp)與小龍蝦(crayfish)找到相應基因。即是從不會飛的甲殼類中找到翅膀基因(pdm和apterous),但是這並非直接的證據,因為動物經常攜帶不表現的基因(可能為祖先原有但現今不表現的基因)。
- 此外,學者設計能夠將pdm和apterous基因衍生的蛋白質染色的化學物質。當他們將此化學物質注入蝦子幼體 體內後,在發育肢體(limb)的上部出現黑色染跡,而那部分則將會發育為鰓(Michalis Averof & Stephen M. Cohen,1997.)[5]。
- 有人對於此假說抱持質疑的態度,因為若此假說成立,則代表昆蟲翅膀由水生昆蟲演化而來,但是在演化學上登陸陸地的現象不會出現兩次(第一次為無翅昆蟲,第二次為有翅昆蟲)(Gullan, PJ & Cranston, PS. 2010.)[2]。
- (三) Endite-Exite Hypothesis:
- 在1978年Kukalova-Peck第一次提出的理論為翅膀由原始昆蟲側體面的外部腿節(exites)演化而來(Kukalova-Peck, J. 1982.)[6],而在1989年Trueman延伸此假說認為應當包括內部腿節(endites),他認為在演化成翅膀的過程中,外部與內部腿節逐漸融合(Trueman, JWH. 1989.)[7]。而此假說具有化石與分子學的證據支持(Jockusch, EL & Nagy, LM. 1997.)[8]
- 三、結論:
- 以上三種昆蟲翅膀的假說皆具有各自支持與質疑的論點,然而光從分子學分析與化石證據中所得出的假說卻未必是昆蟲翅膀真正的演化過程,因為仍然有許多當代因素是現今人類無法完全得知的,這也是為何目前假說所無法完全解釋的原因之一。
- 四、參考文獻:
- [1]Wootton, RJ. 1976. In: Insect flight Symposia of the Royal Entomological Society of London(eds Rainey, RC) p. 235 - 254. Blackwell Scientific Publications
- [2] Gullan, PJ & Cranston, PS. 2010. The insects: and outline of entomology. Wiley-Blackwell, United Kingdom
- [3]Grimaldi, David; Engel, Michael S. 2005.Evolution of the Insects. New York, NY: Cambridge University Press
- [4]Woodworth, CW. 1906. The wing veins of insects. University of California Publishing. Technical Bulletin. Entomology 1, 1 – 52
- [5]Michalis Averof & Stephen M. Cohen,1997,Nature 385, 627 – 630
- [6]Kukalova-Peck, J. 1982. Origin of the insect wing and wing articulation from the arthropodan leg. Canadian Journal of Zoology 61, 1618 – 1669
- [7] Trueman, JWH. 1989. Evolution of insect wings: a limb exite plus endite model. Canadian Journal of Zoology 68, 1333 - 1335.
- [8]Jockusch, EL & Nagy, LM. 1997. Insect evolution: How did insect wings originate Current Biology 7, R358 - R361
andrewneteye4343 發表在 痞客邦 留言(0) 人氣()
昆蟲頭部的體節發育
- 一、前言:
- 現今關於昆蟲頭部體節的假說多樣,從五體節假說到七體節假說都有人提出來,主要是藉由胚胎發育、分子鑑定及形態比較等方式推測頭部體節的起源與演化,有人推測在五百萬年前海洋泥層中兩側對生動物的可能始祖Urbilaterian(Knoll, AH, Carroll, SB (1999))[1],可能已出現獨立於身軀的頭部(De Robertis, EM, Sasai, Y (1996))[2],但因為缺乏化石證據使得此假說無法被大眾廣為接受。在談論頭部前,應當了解頭部確實的定義,頭部指動物前部末端的複合式結構,通常有頸部區分頭部與身軀,頭部包括感覺與神經中心,透過研究頭部功能性與形態結構可以追溯頭部的演化史。
- 二、正文:
- 所有兩側對生動物的身軀由Hox基因群(Hox cluster)調控分區,而不同物種的Hox基因群有不同的組成與表現程度,這使得物種間的身軀分區不盡相同(Kmita, M, Duboule, D (2003))[3];而頭部前段則是由自由Hox基因表現區域(free Hox gene expression zone),此段區域由Orthodenticle(一種轉錄因子)調控,在脊椎動物與節肢動物間有高度保守性(Reichert, H, Simeone, A (1999))[4],因此針對Orthodenticle進行分析時可以從微小變化推測物種間的差異,也可以做為分類物種的參考依據之一,但儘管知道Orthodenticle在系統發育上有高度保守性,目前對於頭部基礎分區化的機制的了解有限。
- 就形態學而言,昆蟲頭部由多個體節組成,目前有多種體節假說,六體節假說為將頭部分為上唇(labrum)、觸角(antennae)、後觸角、大顎(mandibles)、小顎(maxillae)及下唇(labium)(P.J Gullan&P.S Cranston(2004)[5]。五體節假說則是將頭部分為觸角、intercalary(為第二對觸角或第二對觸角退化處)、下顎(mandibular)、上顎(maxillary)及唇(labial)。 儘管有多種頭部體節假說,但是因為頭部前部(anterior head region)缺乏與身軀明確的形態相似性,可能原因為頭部結構有高度適應性,而利於快速演化,使得祖先相似性逐漸模糊而難以被察覺。
- 在胚胎發育學中,難以採用果蠅作為模式動物觀察頭部的發育,原因是果蠅在胚胎發育過程中,胚胎前端會被拉進胚胎內,因此難以觀察到實際的頭部發育過程。基於上述理由,學者認為赤擬穀盜(Tribolium castaneum)可以作為節肢動物胚胎時期頭部發育的模式動物,原因為赤擬穀盜幼蟲具有典型的昆蟲頭部結構,在實驗室環境下全年皆可大量生產子代,最後赤擬穀盜適合進行遺傳學上正向或是負向調控的實驗 (Klingler, M (2004))[6]。 利用非物種特異性轉化系統(non-species-specific transformation system),而赤擬穀盜在廣範圍轉位子基因轉移促進基因轉移的過程中具有高效率(Berghammer, AJ, Klingler, M, Wimmer, EA (1999)[7],這項技術成功地運用於分析增強子(enhancers)(Eckert, C, Aranda, M, Wolff, C, Tautz, D (2004))[8]並產生利於插入突變的物質,(Horn, C, Offen, N, Nystedt, S, Hacker, U, Wimmer, EA (2003))[9]。
- 赤擬穀盜具有強大的逆向基因性(reverse genetics),使得可與RNAi高度結合(Brown, SJ, Mahaffey, JP, Lorenzen, MD, Denell, RE, Mahaffey, JW (1999))[10],藉此可以快速進行染色體的分離與功能性分析。藉此比對不同節肢動物的頭部發育過程,可以協助學者了解頭部發育機制的演化保守性與適應環境的現象。
- 三、結論:
- 從初步實驗結果看來,赤擬穀盜頭部發育間隙 (gap)與類間隙 (gap-like)基因比果蠅頭部更具有功能性的變化。之後將會採取廣泛的比較方法來確定哪些功能為特異性,哪些為系統發育的保守性。此外,特定結構的形態相似性可能顯示結構的同源性,然而在演化過程中基因的表現可能會導致結構的融合或分離。

-
圖一、比較果蠅與赤擬穀盜頭部發育差異
andrewneteye4343 發表在 痞客邦 留言(0) 人氣()


